
疼痛途径
作者: 医知苑
最后更新时间: 2024-05-02
作者: 医知苑
最后更新时间: 2024-05-02
疼痛是一种身体和情绪上的感觉,本质上是令人不愉快的,并且与实际或潜在的组织损伤有关。从生理学角度来看,疼痛功能对于生存至关重要,并且具有重大的进化优势。引起疼痛的行为通常是危险且有害的。这意味着它们通常不会被加固和重复,从而减少造成损坏的可能性。
疼痛的分类很复杂。有许多不同类型的疼痛,每种疼痛都是通过独特的机制产生的。疼痛的类型包括:锐痛、刺痛、热痛或酸痛。疼痛的来源可以是躯体性疼痛、内脏性疼痛、丘脑性疼痛、神经性疼痛、心身性疼痛、牵涉性疼痛或幻觉性疼痛。疼痛本质上也可以是急性或慢性的。
本文将概述疼痛的“经典”图景,即当我们绊倒脚趾或触摸尖锐物体时所感受到的疼痛。它将重点关注疼痛通路如何在脊髓内启动和处理。
一般疼痛途径
在疼痛通路内,有 3 个神经元序列携带发出疼痛信号的动作电位:
- 一阶神经元——这些是伪单极神经元,其细胞体位于背根神经节内。它们有一个轴突,分裂成两个分支,一个外周分支(向外延伸)和一个中央分支(从中央延伸到脊髓/脑干)。
- 二阶神经元——这些神经元的细胞体存在于脊髓的雷克斯德层或脑干内的颅神经的核中。然后这些神经元在脊髓的前白色连合中交叉,并在脊髓丘脑束中向上上升到丘脑的腹侧后外侧(VPL)核。
- 三级神经元– 三级神经元的细胞体位于丘脑的VPL内。它们通过内囊的后肢投射,终止于同侧中央后回(初级体感皮层)。中央后回是按躯体组织的。因此,手部发出的疼痛信号将终止于专门负责手部感觉的皮层区域。
下面是疼痛通路的简单概括,但是更详细的模型超出了本文的范围。
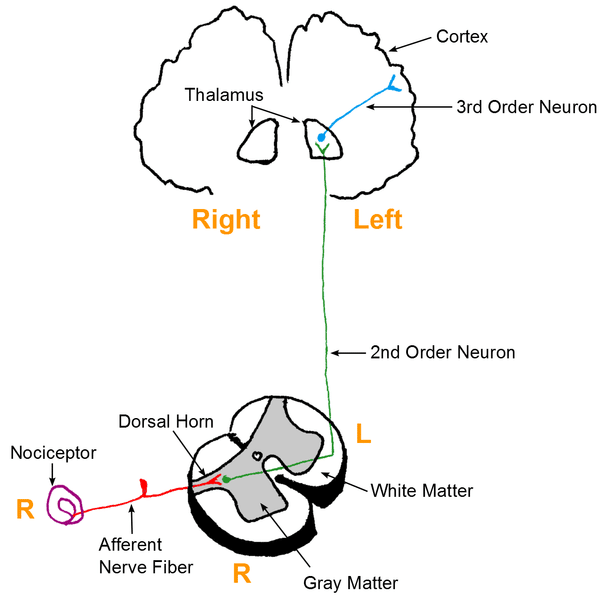
图 1 – 显示伤害感受的简化一般路径的图表。
一阶神经元的激活
伤害感受器
一些一级神经元具有称为伤害感受器的特殊受体,它们通过各种有害刺激被激活。伤害感受器存在于初级传入神经元的游离神经末梢。
由于伤害感受器是游离的神经末梢,这意味着它们是未封装的 皮肤感受器。这与封装的皮肤感受器(例如默克尔盘)相反,后者检测其他感觉模式,例如皮肤的振动和拉伸。
与其他感觉方式类似,每个伤害感受器都有自己的感受野。这意味着当皮肤的特定区域受到刺激时,一个伤害感受器将传导疼痛信号。整个身体的感受野的大小各不相同,并且经常与邻近的感受野重叠。
指尖等区域的感受野比前臂等区域小。此外,它们在该感受野内具有更大密度的游离神经末梢。这种差异很重要,因为它可以提高检测感官刺激的敏锐度。
特定身体部位的体感皮层中皮质表征的大小与该身体部位的感受野的大小相关。例如,由于指尖的感受野较小,因此感觉敏锐度较高,因此它们具有较大的皮质表征。
伤害感受器存在于皮肤、肌肉、关节、骨骼和器官(大脑除外)中,并且可以对许多不同的刺激做出反应。存在不同类型的伤害感受器:
- 机械伤害感受器——检测皮肤的扩张(拉伸)和压力,从而引起尖锐的刺痛。
- 化学伤害感受器——检测外源性和内源性化学制剂,例如前列腺素、组胺等。
- 热和机械热伤害感受器——检测引起缓慢灼痛或冷而剧烈疼痛的热感觉。
- 多模式伤害感受器——检测机械、热和化学刺激。
传输至脊髓
来自机械、化学、热和机械热伤害感受器的信号主要通过Aδ 纤维传输到脊髓背角 。这些有髓纤维的发射阈值低,传导速度快。因此,它们负责传递最初感受到的疼痛。
此外,Aδ 纤维允许疼痛定位并形成疼痛引起的反射的传入通路。 Aδ 纤维主要终止于Rexed laminae I,在那里它们主要释放神经递质谷氨酸。
多模式伤害感受器通过C 纤维将信号传输到背角。 C纤维无髓鞘,传导速度慢。因此,C 纤维导致了我们感受到的继发性疼痛,这种疼痛本质上通常是钝痛、深痛和搏动性疼痛。这些纤维通常具有较大的感受野,因此导致疼痛定位不良。
与Aδ纤维相比,C纤维的点火阈值较高。然而,有害刺激会导致 C 纤维敏感并降低其放电阈值。 C 纤维主要终止于Rexed laminae II(称为凝胶质)并释放神经递质物质 P。其他神经递质由终止于脊髓内的初级传入神经元释放,例如天冬氨酸和血管活性肽。
组织损伤后会释放多种因子,导致伤害感受器激活。这些包括:
- 花生四烯酸
- 钾
- 5-羟色胺
- 组胺
- 缓激肽
- 乳酸
- ATP
其中许多因素也是促炎性的,并导致损伤区域发生急性炎症。
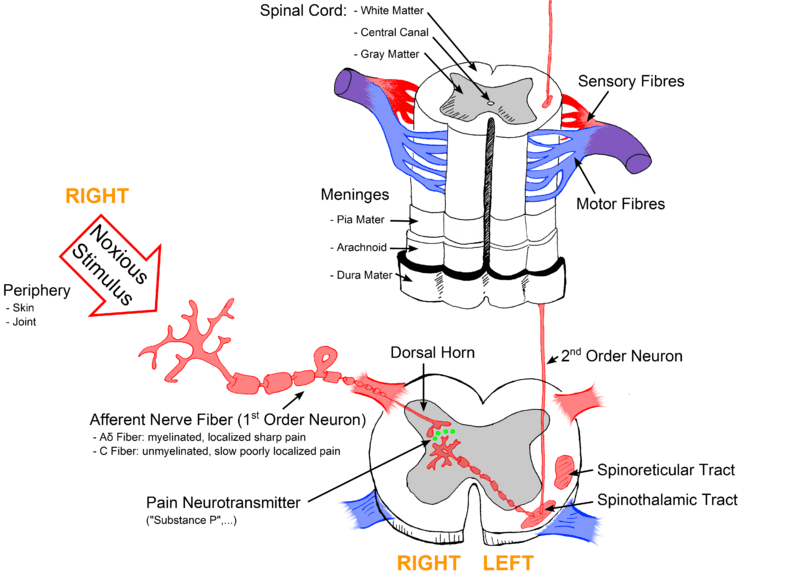
图 2 – 该图更详细地表示了疼痛通路,显示了背角内不同的传入纤维和神经递质释放。
对疼痛的感知改变
痛觉过敏
痛觉过敏描述了在正常阈值刺激下疼痛感增强的现象。痛觉过敏的病理生理学被认为是由受损区域及其周围的神经敏化引起的。 敏化意味着这些神经因有害刺激而降低了放电阈值。外周致敏的一个例子是游离神经末梢释放 P 物质,刺激周围细胞释放分子,从而加剧疼痛。
致敏也可能发生在脊髓中央。病理生理学被认为是由于NMDA受体数量的增加以及 NMDA 受体对谷氨酸敏感性的增加引起的。这些变化发生在二级神经元的树突上,是一级神经元延长谷氨酸释放的结果(由于持续的伤害感受器激活)。
中枢敏化导致脊髓背角内二级神经元过度兴奋。
异常性疼痛
异常性疼痛是对以前不痛的刺激做出反应而产生的疼痛感。在受有害刺激影响的区域及其周围也可以观察到这种情况。
痛觉过敏和异常性疼痛是对疼痛的生理反应的特征,但也可以在病理学中观察到。例如,神经性疼痛的特征是痛觉过敏、异常性疼痛和由于神经损伤引起的伤害感受器自发放电。
疼痛的递减调节
在中枢神经系统内,存在三种类型的阿片受体调节疼痛信号的神经传递。这些受体称为 mu、delta 和 kappa 阿片受体。
它们都是G 蛋白偶联受体,它们的激活会导致神经递质释放减少和细胞超极化,从而降低细胞兴奋性。外源性阿片类药物,例如吗啡,通过作用于这些受体提供良好的镇痛作用。同样,我们的身体含有内源性阿片类药物,可以在生理上调节疼痛。内源性阿片类药物分为三种类型:
- β-内啡肽– 主要与 mu 阿片受体结合
- 强啡肽– 主要与 kappa 阿片受体结合
- 脑啡肽——主要与 δ 阿片受体结合
阿片类药物可以在多个层面上调节疼痛,包括脊髓、脑干和皮质。在脊髓内,强啡肽和脑啡肽都可以起到减少背角疼痛信号传递的作用。这是因为二阶神经元的突触后末端膜内有阿片受体。此外,一级神经元的突触前末端含有阿片受体。
当内源性阿片类药物作用于这些受体时,它会减少一级神经元释放的神经递质,并引起二级神经元的超极化。总之,这减少了二阶神经元动作电位的激发,阻止了疼痛信号的传递。
临床相关性 -世界卫生组织 (WHO) 镇痛梯
世界卫生组织制定了控制疼痛的逐步流程。这个梯子有 3 个台阶。如果第一步没有控制疼痛,患者将继续下一步。
- 第 1 步(轻度至中度疼痛) –非阿片类药物(扑热息痛、阿司匹林或 NSAID)+/- 佐剂(低剂量三环类抗抑郁药/抗惊厥药/肌肉松弛剂/其他 NSAID)
- 第 2 步(中度至重度疼痛) –弱阿片类药物(可待因/曲马多)+/- 非阿片类药物 +/- 佐剂
- 第 3 步(剧烈疼痛) –强阿片类药物(吗啡/芬太尼/二吗啡)+/- 非阿片类药物 +/- 佐剂